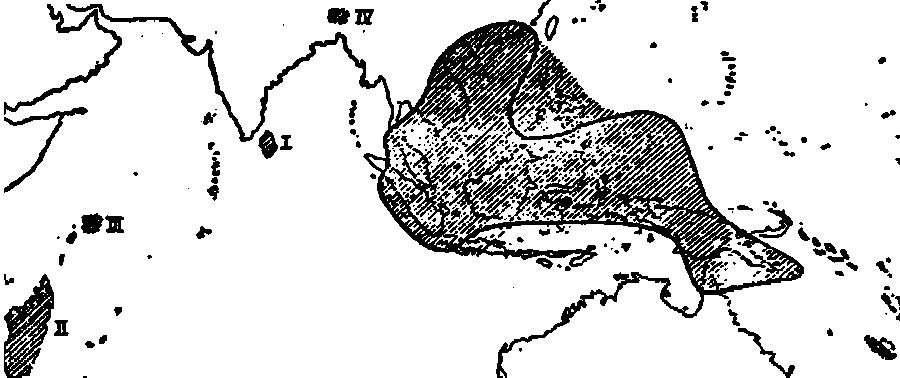
 Fig. 27. Hatched: the total area of Nepenthes ; isolated in the western
part. I. N. destillatoria ; II. N. madagascariensis ; III. N.
Pervillei ; IV. N. khasiana ; enclosed by a line ; N.
mirabilis.
Fig. 27. Hatched: the total area of Nepenthes ; isolated in the western
part. I. N. destillatoria ; II. N. madagascariensis ; III. N.
Pervillei ; IV. N. khasiana ; enclosed by a line ; N.
mirabilis.GENERAL DISCUSSIONS.
1: Phytogeographical.
The total area of distribution of the genus Nepenthes extends westwards and southwards to Madagascar, eastwards and southwards to the Isle of Pines, southeast of New Caledonia, northwards to the Khasi Mountains and southern China. On the mainland of Africa and in America Nepenthes does not occur and in Australia it is only found in the Cape York Peninsula. So the distribution from East to West covers a much greater distance than that from North to South. This is not at all surprising. Nepenthes thrives in wet tropical countries and it is for that very reason remarkable, that it does not occur farther east- and westwards. At first sight one is inclined to think, that the dispersion to the east is limited by the Pacific, but we will perceive, that this is not quite so. There seems to be nothing to hinder the dispersion towards Africa, so that provisarily it must be accepted, that the limited distribution of Nepenthes is due to the relative youth of the genus. Subsequent facts will tend to corroborate this supposition ; the theory of WEGENER, however, throws new light on this question, as we will see in the discussion of the phylogeny of this genus.
Considering more in detail the limits of the area and starting with Sumatra, we may remark firstly, that, since we know that several species have already been found on the islands off the western coast the dispersion has been limited here by the Indian Ocean. It seems strange, that the boundary cuts off the northern part of Sumatra, but taking into consideration, that these regions are very little known from a floristic point of view, and that north of Lake Toba new species have been discovered during the last few years, we may readily suggest, that after a more careful examination of the country. at least some widely spread species will prove to occur there.
.
Fig. 27. Hatched: the total area of Nepenthes ; isolated in the western
part. I. N. destillatoria ; II. N. madagascariensis ; III. N.
Pervillei ; IV. N. khasiana ; enclosed by a line ; N.
mirabilis.
Following the boundary we observe it running west and south of Sumatra and Java, crossing the latter island at about 4/5 of its length, continuing along the coast of Borneo, cutting off the southern part of the southwestern peninsula of Selébès, running south of the Moluccas and along the coast of New Guinea, including only the north-eastern peninsula of Australia, finally bending northwards south of the Isle of Pines it is obvious, that the exclusion of the Lesser Sunda Islands and the southern part of Selébès is due to climatic conditions, since these regions have, under the influence of the mainland of Australia, to endure a dry season, lasting too long for the existence of Nepenthes.
Looking at the eastern limit of the area we are struck by the fact, that from the islands north and east of New Guinea only the Palau Islands possess Nepenthes and then only one species. The Bismarck Archipelago, the Solomon Islands and the Fiji Islands are sufficiently exploited, so that we may therefore accept, that Nepenthes does not occur there. The climate of these islands, however, is certainly not less favourable for Nepenthes than that of New Caledonia, whence we may conclude, that here also the area is apparently limited by the relative youth of the genus.
Following the boundary to the North we see, that Nepenthes is spread throughout the Philippine Islands, but that it has not reached Formosa, though it occurs in southern China at the same latitude. The course of the boundary on the Asiatic continent is not exactly known. Nepenthes has been found in different parts of Farther India and in the Malay Peninsula south of the isthmus, but not along the Gulf of Siam (where it undoubtedly occurs), nor in Burma or on the coast north of it, but it is found in the Khasi Mountains. lt is doubtful, whether the isolated occurrence of Nepenthes in the Khasi Mountains is actually a case of discontinuous distribution or is merely a result of the lack of knowledge regarding the flora of parts of Farther India, where Nepenthes may be eventually discovered.
Nepenthes has not been found in the Andaman and Nicobar islands, but it occurs in Ceylon, the Seychelles and Madagascar. Apparently the genus has already found the difficult way to Madagascar and if this suggestion is right, it seems probable that in future it will also be able to reach Africa.
We now pass to the discussion of the areas of the separate species. Of the 65 species, acknowledged by me, 4 (N. destillatoria, N. alata, N. Pervillei and N. madagascariensis) are isolated in the western end of the generic range, each of them covering an area of its own and not occurring along with any other species (cf. fig. 27). This peculiar fact I shall have to consider again when discussing the phylogeny. Another remarkable fact is, that so many species have been found in one spot or at least on 2 single mountain only, viz. 25, of which 13 occur in Borneo, 2 in Mindanao, 2 in Palawan, 2 in Cochin China and 3 in New Guinea. On the maps these species have been indicated by dots. (cf. fig. 28 to 34.)
The remaining 36 species occupy very different areas. When we look at the species with wide distributions, we should mention in the first place N. mirabilis, the range of which covers by far the largest part of that of the whole genus (cf. fig. 27). Westwards this species has spread to Sumatra and the islands west of it, in the Philippines it has reached only as far as Mindanao, in the Southeast it extends as far as the Louisiade Archipelago, in Java it occurs in the western quarter only. Another species of wide range is N. ampullaria (cf. fig. 33), which occupies two separate but widely distant areas: one on the Sunda-shelf and one in New Guinea. As N. ampullaria prefers the coastal regions it is impossible to ascribe this to the insufficient floristic exploration of Selébès and the Moluccas. The third species with a wide range is N. maxima (fig. 32), which is distributed from Borneo to New Guinea. It is not clear, why this species has not spread farther to the west.
Passing now to the 33 species with average-sized areas, we shall discuss in the first place those, which occupy areas more or less congruent with the Sunda-shelf, i.e. the western part of the Archipelago, including Sumatra, the Malay Peninsula, Borneo and Java. Six species have such: N. albo-marginata, N. gracilis, N. Hookeriana, N. Rafflesiana, N. Reinwardtiana and N. tentaculata (fig. 28, 33 and 34). Of these, N. Hookeriana needs no consideration, as it is probably a hybrid of N. Rafflesiana and N. ampullaria. The total area of the first and the western part of the area of the latter being also congruent with the Sunda-shelf. Of the other 5, N. tentaculata occurs almost exclusively in Borneo, having been collected only once in Selébès. The range of N. gracilis almost exactly covers the Sunda-shelf ; it has, however, been found twice in Selébès. In the case of N. Rafflesiana, N. Reinwardtiana and N. albo-marginata the congruence of their area with the Sunda-shelf is equally almost complete. Undoubtedly this has a geological cause, since the Sunda-shelf has, during the Tertiary Era, been alternately dry and partly flooded by shallow seas, whereas its eastern boundary is formed by the Straits of Makassar, for which a great age is generally accepted.
Within the Sunda-shelf we may distinguish several smaller areas, that some species seem to prefer. On fig. 30 we remark in the first place 4 circles: in the Malay Peninsula, Sumatra, Java and southern Borneo, the latter 3 of which being united by lines. The circles indicate the areas of 10 species, all nearly related, 3 of which occur in the Malay Peninsula and nowhere else (viz. N. Macfarlanei, N. gracillima and N. sanguinea), 6 in the mountains of central Sumatra only (N. Bongso, N. carunculata, N. dubia, N. inermis, N. pectinata and N. singalana), and one in the last named region, in Java and on one mountain in southern Borneo (N. gymnamphora). Of these 10 species N. dubia belongs to those, that have been found on one mountain only.
Another peculiar region is that in the mountains of northwestern Borneo (fig. 31, 32 and 34) to which 5 species are limited: N. bicalcarata, N. hirsuta, N. Lowii, N. stenophylla and N. Veitchii, whereas 7 other species are found in this region on only one mountain. Perhaps this too has a geological significance.
Other remarkable areas within the Sunda-shelf are that of N. tobaica, north, east and south of Lake Toba, and that of N. alata, which inhabit Central Sumatra, the Malay Peninsula, the Philippine Islands, perhaps as far as Palawan, but which has not yet been discovered in Borneo.
Looking now at New Guinea (fig. 29 and 33), we may observe 4 species in the mountains of this island, occurring in a region apparently rich in Nepenthes, viz. N. neoguineensis, N. papuana and N. insigne which are confined to this area, and N. Vieillardii, which occurs also in New Caledonia and in the Isle of Pines. Moreover, 3 species have been found in the same region on a single mountain only.
Finally a few words regarding the remaining species from the Philippine Islands and Cochin China. On the highest mountains of Mindanao there occurs, besides the previously mentioned species that are found on one mountain only, also N. Merrilliana, perhaps also collected once in Selébès, whereas on the islands north of Mindanao occur N. Burkei and N. ventricosa, which are perhaps mere forms of a single species (cf. fig. 33). In Cochin China (fig. 28) there are 2 species found in one locality only and 2 species occupying larger areas. Our knowledge of the Cochin China Nepenthes, however, is mainly due to the investigations of the last two decades and is still very incomplete.
The regularities discussed, though insignificant of themselves, have a greater value for the discussion of the phylogeny.
2. Taxonomical.
Not only from a morphological and physiological, but also from a taxonomical point of view, Nepenthes is a remarkable genus. Whereas the numerous species are mutually so nearly related, that the separation is often very difficult, the genus stands quite isolated among all other dicotyledoneous plants. The relations have been sought in two directions. Ancient systematicians set little value on the vegetative organs and sought the affinities among plants with similar flowers, such as the Cytinaceae and the Aristolochiaceae. Modern taxonomists, for whom the generative organs are not alone, sufficient on which to base a phylogenetic explanation of the system, sought connections with other pitcher plants, such as the Sarraceniaceae and Cephalotaceae. After all, however, these families are so entirely different from the Nepenthaceae, that none of them can be joined with this order in the same division, nor can they be considered as closely related from a phylogenetical standpoint.
As I have already pointed out, the question of the relations between the species is quite another. The first months during which I was studying this genus and investigating the natural limits between the species I often got the impression, that perhaps most of the accepted species might be only the most striking or the most common varieties of a relatively small number of species. Continued studies have proved, that this is not correct. Several species, among them some nearly related ones, soon appeared to be quite sharply distinguishable, such as N. albo-marginata, N. Reinwardtiana, N. gracilis, to say nothing of the much more distinct species, such as N. ampullaria, N. Rafflesiana, N. bicalcarata. And further study showed more and more species to be well defined. It must be admitted, that among the species acknowledged in this revision, several may be wrongly limited, but undoubtedly further studies will rectify these mistakes.
Nevertheless it has not been possible to divide the genus Nepenthes into distinct subgenera. The more or less natural groups which I have distinguished below, perhaps have some phylogenetic value, as may be discussed later on, and we may call them sections.
The combination of the sharp limitation externally and the difficulty of an internal division result in making Nepenthes one of the most natural groups of the vegetable kingdom.
About the limitation of the species, hybridization experiments will be required in order to make a final decision and in the first place the possibilities of the propagation of the hybrids and their relative fertility must be known. Now, in the last century a large number of Nepenthes have been cultivated and hybridized, but this has not resulted in a better idea of the species limits. The hybrids have been put on the market, they have been crossed with other hybrids and species, and the products of these crossings also have been offered for sale, but no exact data on the fertility of the hybrids an the polymorphy of their progeny after inbreeding are to be found in the botanical or horticultural literature. N. Rafflesiana and N. ampullaria, for instance, have often been grown, but their hybrid has never been recorded though this experiment would have given the solution of the question, whether N. Hookeriana is identical with this hybrid. The breeders have also cultivated N. Hookeriana, they have hybridized it with other species and hybrids, but it has never been recorded, whether the resulting progeny was more polymorphous than that of other cross-breedings. It would be of great scientific value, if plant breeders would observe the possibilities of crossings, and also publish the hybridizations which result in failure, observe the grade of fertility of the hybrids and study the polymorphy of the progeny. Regarding the theory of species limits some space will be devoted to the consideration of this subject when discussing the phylogeny of the group.
In the following paragraphs an attempt has been made to work out a natural division of the genus. When studying the species I had already observed, before seeking a systematic arrangement, that groups of related species could be distinguished ; this is a common phenomenon in almost all large genera, but it was the more remarkable in the genus in question, since the distinction of the species was so difficult. When trying to define more precisely this division it proved to be possible to arrange the species into 6 groups, being quite different mutually regarding the number of species. I present the arrangement of the species in a tabulated statement and will discuss it in detail.
The first group is that of the Vulgatae. It comprises the most simple forms, which in general have no coarse stems, no large nor broad, mostly lanceolate leaves, a phyllotaxis almost always 2/5, no hirsute reddish nor brown indumentum, not large and rarely infundibuliform pitchers, a narrow peristome, mostly no, very rarely one appendage on the lower surface of the lid and a paniculate or racemose inflorescence, while the glands of the inner surface of the pitcher are often little or not overarched. The species are distributed all over the generic area.
The species belonging to the second group, that of the Montanae, have in general the same characters, but the plants are usually more robust, the phyllotaxis is always 2/5, the pitchers are often larger, the upper ones often infundibuliform, the peristome is generally broader, the colour of herbarium specimens not rarely blackish ; the leaves are always sessile, the inflorescence is a raceme. They are all closely related to N. gymnamphora, and are found in the mountains of the Malay Peninsula, Sumatra, Java and perhaps in those of southern Borneo.
Synoptic statement of the species.
Vulgatae
N. destillatoria LINN.1) 34. N. papuana DANS. 1. N. alata BLANCO
46. N. tomoriana DANS. N. khasiana HOOK.F.4) ? N. philippinensisMACF.6)
30. N. neoguineensis MACF. 14. N. gracilis KORTH.
48. N. trichocarpa MIQ. 27. N. mirabilis DRUCE
33. N. paniculata DANS. 44. N. tentaculata HOOK. F.
N. madagascariensis 39. N. Reinwardtiana MIQ. 2. N. albo-marginata LOBB
POIRET.2) 45. N. tobaica DANS.
? N. Pervillei BL.3) 50. N. Vieillardii HOOK. F. ? N. anamensis MACF.7)5)
N. Thorelii LEC.5) ? N. Geoffrayi LEC.5)
? N. kampotiana LEC.5)
Montanae
16. N. gymnamphora NEES 15. N. gracillima RIDL. 29. N. inermis DANS.
35. N. pectinata DANS. 40. N. sanguinea LINDL. 11. N. dubia DANS.
5. N. Bongso KORTH. 24. N. Macfarlanei HEMSL.
8. N. carunculata DANS.
41. N. singalana BECC.
Nobiles
42. N. spectabilis DANS. 17. N. hirsuta HOOK. F. 29. N. neglecta MACF.
22. N. leptochila DANS. ? N. Deaniana MACF.8)
Regiae
25. N. maxima NEES 36. N. pilosa DANS. 9. N. clipeata DANS.
32. N. oblanceolata RIDL. 7. N. Burbidgeae BURB. N. truncata MACF.9)
49. N. Veitchii HOOK. F. 38. N. Rajah HOOK.F.
13. N. fusca DANS. 12. N. ephippiata DANS. 6. N. Boschiana KORTH.
43. N. stenophylla MAST.
21. N. Klossii RIDL. 28. N. mollis DANS. 23. N. Lowii HOOK. F.
Insignes
26. N. Merrilliana MACF. 31. N. Northiana HOOK. F. 51. N. villosa HOOK. F..
N. petiolata DANS. 10. N. decurrens MACF.
20. N. insignes DANS.
N. Burkei MAST.10) 47. N. Treubiana WARB. 37. N. Rafflesiana JACK
N. ventricosa BLANCO11)
Urceolatae
4. N. bicalcarata HOOK. F. 3. N. ampullaria JACK (18. N. Hookeriana LINDL.)
1). (dist.) LINN., Sp. pl., ed. 1, II, p. 955 (1753) ; MACF., in ENGL., Pflanzenr., IV, 111, p. 35 (1908).
2). POIR., in LAM., Enc. méth., bot., IV, p. 459 (1796) ; MACF., l.c., p. 59 (1908).
3). BL., Mus., II, p. 10 91852) ; MACF., l.c., p. 31 (1908).
4). HOOK. F., in D.C. Prodr.., XVII, p. 102 (1873) ; MACF., l.c., p. 59 (1908).
5). LEC., Not. syst., I, p. 61-64 (1909) ; Fl. Ind. Ch., V. p. 47-51 (1910).
6). MACF., l.c., p. 43 (1908) ; N. brachycarpa MERR., Phil. Journ. Sc., X, bot., p. 306 (1915) sec. MERR., Enum. Phil., p. 215 (1923).
7). MACF., l.c., p. 39 (1908).
8). MACF., l.c., p. 57 (1908).
9). MACF., Contr. Bot. Lab. Pennsylv., III, p. 209, t. II (1911).
10). (Burkeii) Mast., Gard. Chron., 1889, 2, p. 492 et 566, ic. 69 (1889).
11). BLANCO, Fl. Fil., ed. 1, p. 807 (1837).
The group of the Nobiles comprises forms, which are intermediate between the preceding and the following groups. The inflorescence is always a raceme, the lid has never appendages on the lower surface, the peristome is not broad. The indumentum reminds that of the Regiae. They are found in Sumatra, Palawan and Borneo.
.
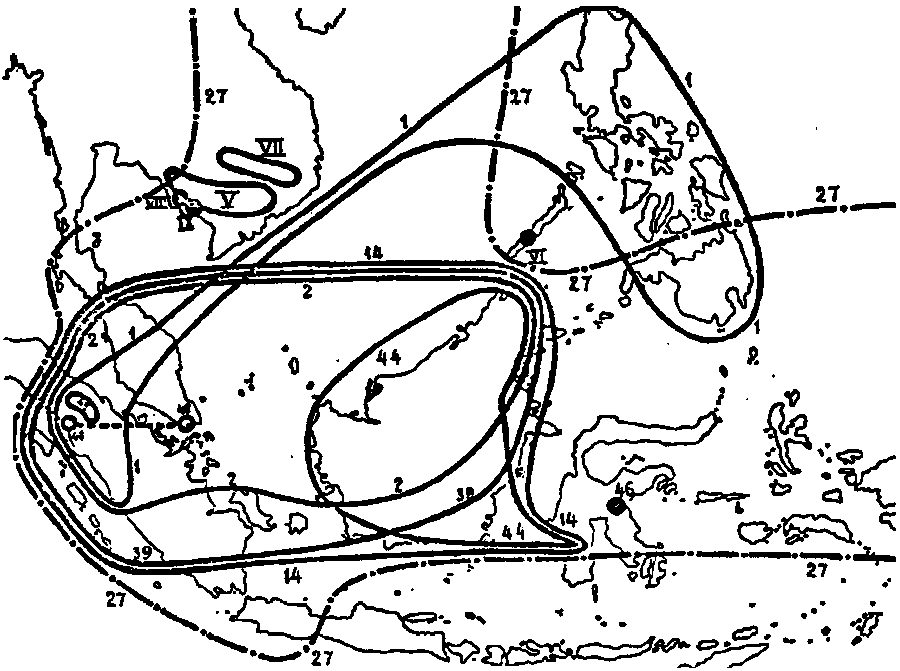 Fig. 28. The distribution of the Vulgatae in the western part of the
Archipelago ; 1. N. alata ; 2. N. albo-marginata ; 14. N.
gracilis ; 27. N. mirabilis (cf. fig. 7) ; 39. N.
Reinwardtiana ; 44. N. tentaculata ; 45. N. tobaica ; 46.
N. tomoriana ; 48. N. trichocarpa ; V. N. Thorelli ; VI.
N. philippinensis ; VII. N. anamensis ; VIII. N. Geoffrayi
; IX. N. kampotiana.
Fig. 28. The distribution of the Vulgatae in the western part of the
Archipelago ; 1. N. alata ; 2. N. albo-marginata ; 14. N.
gracilis ; 27. N. mirabilis (cf. fig. 7) ; 39. N.
Reinwardtiana ; 44. N. tentaculata ; 45. N. tobaica ; 46.
N. tomoriana ; 48. N. trichocarpa ; V. N. Thorelli ; VI.
N. philippinensis ; VII. N. anamensis ; VIII. N. Geoffrayi
; IX. N. kampotiana.
The Regiae comprise the most beautiful and largest species and the most remarkable forms ; In general they are large, coarse plants, with large, petiolate leaves and a coarse, red-brown indumentum ; the phyllotaxis is often 1/2 in the elongated stems ; the upper pitchers are infundibuliform and the lid bears an appendage on the lower surface near the apex. In herbarium specimens the colour is mostly yellowish- or reddish-brown. The peristome is mostly flattened or expanded, the inflorescence is a raceme. The Regiae occur in the first place in Borneo, but in few species they are spread as far as Mindanao and New Guinea.
The fifth group, that of the Insignes, like that of the Regiae, also comprises very beautiful, but in general less aberrant forms. They are large plants with petiolate leaves and bear large pitchers with flattened or expanded peristomes, but the upper pitchers are more campanulate-infundibuliform, the leaves are often sessile, the indumentum is more sparse or even wholly absent and there are never appendages on the lower surface of the lid. The Insignes occur in the first place in Borneo ; few species, however, have been found in Mindanao and New Guinea.
The remaining species form the group of the Urceolatae, similar by the lower pitchers, which are urceolate and glandular over the whole inner surface, and by the peristome, which is flattened at the inner side and very delicately ribbed. The upper pitchers, when developed, are infundibuliform and the inflorescence is a panicle. The phyllotaxis is 3/5. One of the species is confined to the Sunda-shelf, one inhabits the Sunda-shelf and New Guinea, the third is found in northeastern Borneo only.
Considering now each group separately, we first perceive, that among the Vulgatae we may distinguish between the species with a racemose inflorescence and those with a paniculate one. Those with a panicle have been placed in the first column, this character, for the remainder, only occurring in the group of the Urceolatae, while the other species show greater resemblance to the following groups.
.
 Fig. 29. The distribution of the Vulgatae in the eastern part of the
Archipelago ; 27. N. mirabilis (cf. fig. 27) ; 30. N.
neoguineensis ; 33. N. paniculata ; 34. N. papuana ; 50.
N. Vieillardii.
Fig. 29. The distribution of the Vulgatae in the eastern part of the
Archipelago ; 27. N. mirabilis (cf. fig. 27) ; 30. N.
neoguineensis ; 33. N. paniculata ; 34. N. papuana ; 50.
N. Vieillardii.
The species with panicles may be discussed in two groups of three. The first are N. destillatoria, N. tomoriana, and N. neoguineensis. These are found in Ceylon, Selébès and New Guinea respectively, and are so closely related, that it is doubtful, whether they are not forms of one single species. Provisorily they can be distinguished by slight differences in the inflorescence, leaves, nervation, pitchers &c. More distinctly different are the other three species of the first column, viz. N. paniculata, N. madagascariensis and N. Pervillei. Of these I have only examined N. paniculata, to which, according to the description, N. madagascariensis shows great resemblance, the latter being distinguished from the former by several less important characters. N. madagascariensis is nearly related to N. Pervillei but the last-mentioned species seems to be easily distinguishable by the sessile leaves, whereas in the two others the leaves are petiolate.
The Vulgatae of the second column are the most simple representatives of the genus, possessing a racemose inflorescence and narrow, sessile leaves. N. papuana forms the connection with the foregoing species, being easily distinguishable from N. neoguineensis only by the racemose inflorescence. Very similar to N. papuana seems to be N. khasiana, only known to me from description and insufficient herbarium materials, but sufficiently different for specific distinction. More important are the differences with N. gracilis, N. trichocarpa and N. tentaculata. Though N. trichocarpa, has its upper pitchers more or less infundibuliform, it is so nearly related to N. gracilis that its place within this group needs no argument. N. Reinwardtiana and the very closely related N. tobaica show a great resemblance with N. gracilis at first sight, but prove to be quite different from it on further examination. The same may be said of N. Vieillardii and the closely related N. Thorelli.
The third column of the Vulgatae comprises the species, which show greater deviations from the general type of the group in different directions. N. alata possesses a character, which is found in no other Vulgata and which in the genus otherwise occurs only in the Regiae, viz. the appendage on the lower surface of the pitcher. Moreover the phyllotaxis is sometimes 1/2 on the elongated stems. However, the appendage an the underside of the lid is often wanting and the phyllotaxis is an inconstant character, and otherwise N. alata is a real Vulgata in every respect ; like in the following species, the leaves are petiolate. N. mirabilis is somewhat isolated by the distinctly petiolate leaves with many longitudinal nerves, which remind of those of N. bicalcarata, but otherwise this species also is a typical Vulgata. N. albo-marginata is peculiar by the abundant indumentum, the whitish colour and the thickened tomentose zone below the peristome, but for the rest this species too is a real Vulgata. Other species of different groups have also often a tomentose zone below the peristome, but not a thickened nor a whitish one.
The remaining three species: N. anamensis, N. Geoffrayi and N. kampotiana, are Cochin China Nepenthes, only known to me from description and perhaps not belonging to this group.
The group of the Montanae comprises the species which are related to N.
.
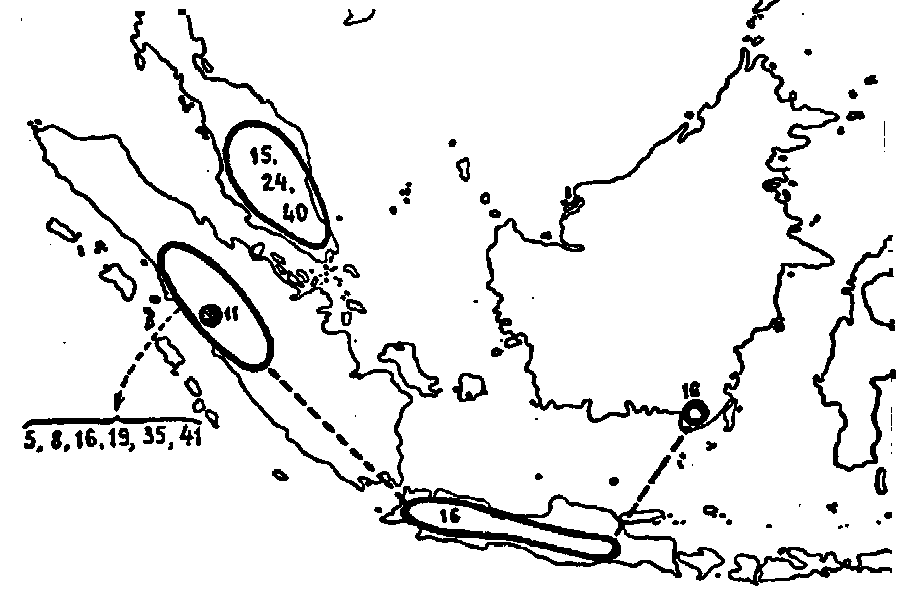 Fig 30. The distribution of the Montanae ; 5. N. Bongso ;
8. N. carunculata ; 11. N. dubia, 15. N. gracillima ; 16.
N. gymnamphora ; 19. N. inermis ; 24. N. Macfarlanei ; 35.
N. pectinata ; 40. N. sanguinea ; 41. N. singalana.
Fig 30. The distribution of the Montanae ; 5. N. Bongso ;
8. N. carunculata ; 11. N. dubia, 15. N. gracillima ; 16.
N. gymnamphora ; 19. N. inermis ; 24. N. Macfarlanei ; 35.
N. pectinata ; 40. N. sanguinea ; 41. N. singalana.
gymnamphora and several of them are connected by intermediate forms, perhaps hybrids.
We start with N. gymnamphora, the most widely dispersed species of this group, and most similar to the Vulgatae. When no other species of this group were known, N. gymnamphora could very well be placed into the preceding one. It would therefore not be so wrong at all to consider the Montanae as a division of the Vulgatae.
The nearest relation of N. gymnamphora is N. pectinata. It is found only in the mountains of Central Sumatra and it grows there along with N. gymnamphora, apparently without intermixing. The difference is only in the pitchers and in the rosettes, the latter being always elongated in N. pectinata. The species nearest related to the latter are N. Bongso and N. singalana, both different in the more slender stems and smaller inflorescences and the entire inner margin of the peristome. I have seen intermediates as well between N. Bongso and N. pectinata as between N. pectinata and N. singalana. Between N. Bongso and N. singalana I have found no intermediates, but when the upper pitchers of N. singalana are somewhat infundibuliform, there remain only a number of minor differences which make it possible to distinguish between these 2 species. N. carunculata is very similar to N. Bongso in the form of its pitchers, especially when the excrescence on the lower side of the lid is wanting. Its inflorescence, however, is almost like that of N. gymnamphora and N. pectinata. Like the foregoing species it is found on high mountains in Sumatra.
A peculiar plant of this group is that which has been called N. Junghuhniana by MACFARLANE and inserted by me under the formula N. sanguinea x singalana. It forms a transition to the species of the Malay Peninsula. Some specimens of N. pectinata (especially BÜNNEMEIJER 4028) and also of N. gymnamphora show resemblance with N. sanguinea, viz. in those cases in which the pitchers are extraordinarily large and red and the peristome is broad and expanded, but these plants can not be considered as transition forms. The plant of JUNGHUHN, however, is more similar to N. sanguinea than any other from Java or Sumatra.
N. sanguinea, from the Malay Peninsula, is very well characterized in its typical forms, but it is vaguely limited against N. gracillima and still more vaguely against N. Macfarlanei, often growing along with it. The very striking hairs on the lower side of the lid of N. Macfarlanei are very different in length and in number in different specimens, and short hairs on the underside of the lid occur also often in N. gracillima and N. sanguinea. Also the upper pitchers of N. Macfarlanei are not always widely infundibuliform, whereas those of N. gracillima and especially those of N. sanguinea are often more infundibuliform than is generally accepted. Moreover, the herbarium materials are, in many cases, very incomplete and this makes the determination of the specimens and the stating of the species limits still more difficult. See under N. gracillima x Macfarlanei and under N. Macfarlanei x sanguinea.
To the group of the Montanae I have brought also N. inermis and N. dubia, two new forms from the mountains of Central Sumatra, at first sight very different from the species mentioned above in the form of the pitchers. N. inermis is remarkable for its very narrow lid and the absence of a peristome. N. Lowii, however, one of the Regiae, has also no peristome whereas a narrow lid is likewise found in N. ampullaria, one of the Urceolatae. Consequently, these characters seem to be of little value for the distinction of groups in general. In all other characters N. inermis and N. dubia agree perfectly well with other Montanae, especially with N. Bongso ; N. dubia is apparently a form intermediate between N. inermis and N. Bongso and may be a hybrid.
The third group, that of the Nobiles, comprises the species that form a transition between the Vulgatae and the Montanae on one hand and the Regiae on the other. N. spectabilis, from Sumatra, reminds one of N. sanguinea in many respects but shows a resemblance with the Regiae by the yellowish colour of herbarium specimens and by the red-brown indumentum. N. hirsuta agrees with the Regiae also in colour and indumentum, but herbarium materials without inflorescences are so similar to N. gymnamphora, that they may be confounded with it. N. leptochila is still more closely related to N. gymnamphora, being still less coarse in all parts, and less hairy. As far as may be concluded from the
.
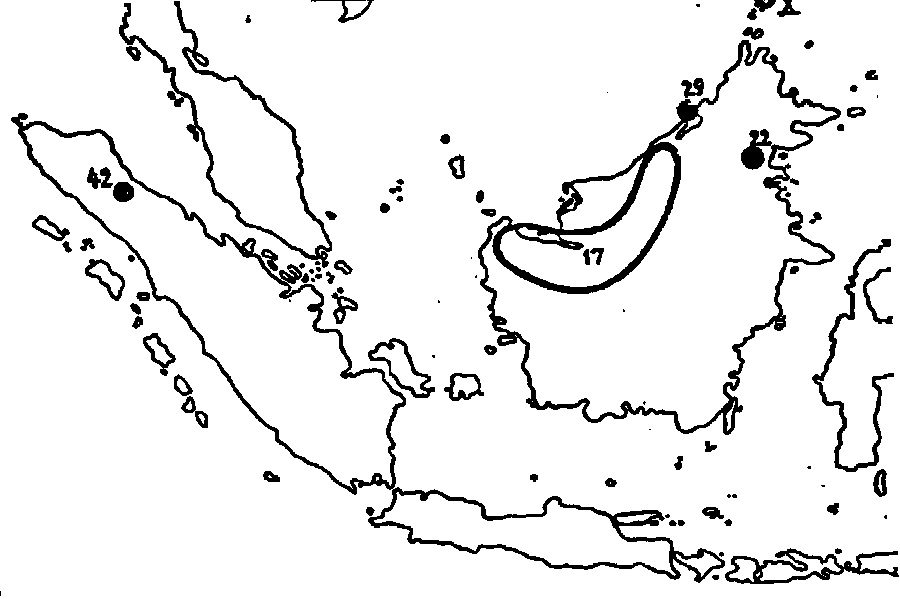 Fig. 31 The distribution of the Nobiles 17. N. hirsuta ; 22.
N. leptochila ; 29. N. neglecta ; 42: N. spectabilis ; X.
N. Deaniana.
Fig. 31 The distribution of the Nobiles 17. N. hirsuta ; 22.
N. leptochila ; 29. N. neglecta ; 42: N. spectabilis ; X.
N. Deaniana.
description, the same may be said of N. neglecta. Less clear is the position of N. Deaniana, which is inserted here provisorily.
We now come to speak of the most striking group, the Regiae. We may start with N. maxima, which is most widely distributed, and best known as to its polymorphy. Though it is more sparsely hairy than most members of its group, it is quite a typical Regia by the the appendages on the lower surface of the lid. Also the mostly infundibuliform upper pitchers, the often broad peristome and the yellowish colour induce us to place it here. N. oblanceolata is placed close after it, as, I presume that it is only an aberrant form of N. maxima. Very closely related, but certainly distinct species are in the first place N. Veitchii, N. stenophylla, N. Klossii and N. fusca. N. Veitchii seems to be almost a mountain form of N. maxima and has already been discussed in detail. Like the other species mentioned it is more typically a Regia than N. maxima by the more dense red-brown indumentum. Less related to the species of the first column, but still typical Regiae, are N. pilosa and the similar N. Burbidgeae, both insufficiently known. The taxonomic place of N. ephippiata and N. mollis is uncertain, as the pitchers are unknown, but the coarse stems and leaves, the yellowish colour of the former and the abundant hirsute indumentum and red-brown colour of the latter leave hardly any doubt whether both are Regiae.
.
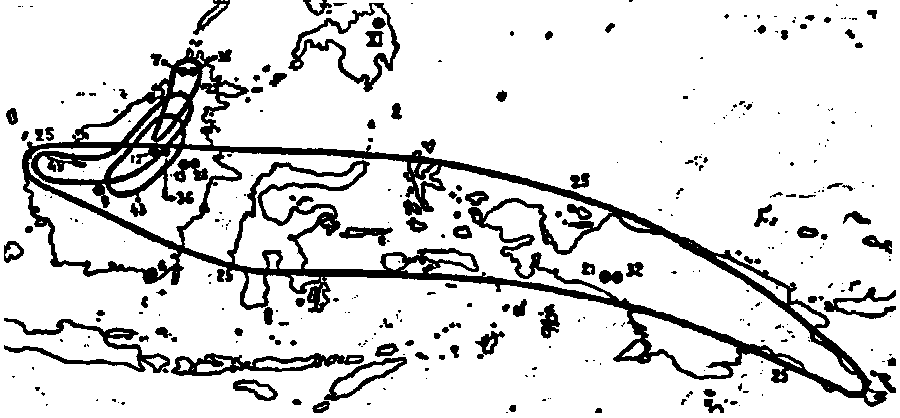 Fig. 32. The distribution of the Regiae ; 3. N. Boschiana ; 7.
N. Burbidgeae ; 9. N. clipeata ; 12. N. fusca ; 21. N.
Klossii ; 23. N. Lowii ; 25. N. maxima ; 27. N. mollis
; 32. N. oblanceolata ; 36. N. pilosa ; 38. N. Rajah ; 43.
N. stenophylla ; 49. N. Veitchii ; XI. N. truncata.
Fig. 32. The distribution of the Regiae ; 3. N. Boschiana ; 7.
N. Burbidgeae ; 9. N. clipeata ; 12. N. fusca ; 21. N.
Klossii ; 23. N. Lowii ; 25. N. maxima ; 27. N. mollis
; 32. N. oblanceolata ; 36. N. pilosa ; 38. N. Rajah ; 43.
N. stenophylla ; 49. N. Veitchii ; XI. N. truncata.
The Regiae of the third column are each peculiar in a different way. N. clipeata has suborbicular leaves, tendrils inserted almost in the middle of the lamina, pitchers consisting of 2 sharply distinguished parts, a subglobose lower part and an infundibuliform upper part, and a vaulted lid. This shape of the pitcher we meet with also in N. ventricosa, of the Insignes group, the vaulted lid also in N. Rafflesiana of the same group. N. truncata has leaves as peculiar as those of N. clipeata, but it shows a resemblance to the Insignes, especially to N. Merrilliana and its allies, by the lack of indumentum and the wide pitchers ; by its lid, however, it belongs to the Regiae. The same may be said of N. Rajah, which has an abnormal leaf form, but an extremely large lid, N. Boschiana is insufficiently known. The infundibuliform upper pitchers, mentioned by KORTHALS, are absent in the type specimens and the indumentum is very sparse, but the broad peristomes and the round lids with appendage place it in this group.
Most abberant is N. Lowii, the leaves and the stem of which are coarse, whereas the indumentum is almost absent and the pitchers show a peculiar form and have no peristome, the lid is vaulted, the midrib is keeled but has no appendage, the lower surface is covered with thick hairs, the glands of the inner surface of the pitcher are so large, that the interspaces are reduced to lines. All these characters, however seem to have little taxonomic value. The form of the pitcher is analogous to that of N. inermis of the Montanae group, which also has no peristome. The peculiar bristles on the lower surface of the lid are found less developed in N. Macfarlanei. The large, flat glands on the inner surface of the pitchers are also found in the lower part of the pitchers of N. Rajah. This is the reason why I have not distinguished a separate group for this species.
.
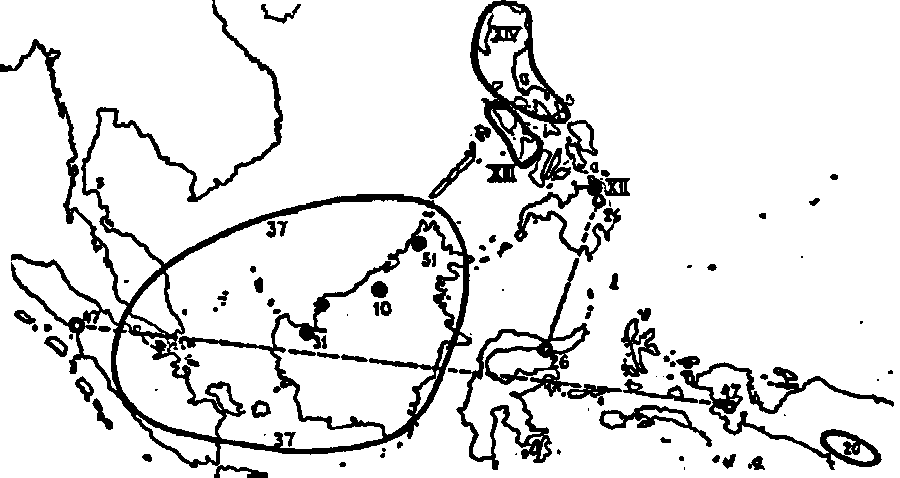 Fig. 33. The distribution of the Insignes ; 10. N. decurrens ;
20. N. insignis ; 26. N. Merrilliana ; 31. N. Northiana ;
37. N. Rafflesiana ; 47. N. Treubiana ; 51. N. villosa ;
XII. N. petiolata ; XIII. N. Burkei ; XIV. N.
ventricosa.
Fig. 33. The distribution of the Insignes ; 10. N. decurrens ;
20. N. insignis ; 26. N. Merrilliana ; 31. N. Northiana ;
37. N. Rafflesiana ; 47. N. Treubiana ; 51. N. villosa ;
XII. N. petiolata ; XIII. N. Burkei ; XIV. N.
ventricosa.
The Insignes are not distinctly separated from the Regiae. Most
striking are N. Merrilliana and the other species of the first column.
They have the
coarse stems with large leaves and pitchers of the
Regiae combined with the slight indumentum, the sessile leaves and lids
without appendages of other groups the species showing this in the same
striking manner as N. Merrilliana are N. petiolata and N.
insignis, whereas N. ventricosa and N. Burkei are somewhat
different. It is not certain whether the latter two species should be
considered as separate species or as forms of a single species. Another type is
formed by the closely related N. Northiana and N. decurrens.
Their upper pitchers are more slender and more infundibuliform, and their
leaves are indistinctly petiolate, but in other respects they are typical
Insignes. N. Treubiana is somewhat isolated by the very
infundibuliform upper pitchers and petiolate leaves ; N. villosa and
N. Rafflesiana are still more isolated, the former by the distinctly
petiolate leaves and the villose indumentum in which it resembles the
Regiae, the latter by the petiolate leaves, the very infundibuliform
upper pitchers and the vaulted pitcher lid.
The remaining species form the group of the Urceolatae, though not showing a strong likeness, yet agreeing by the pitchers and the paniculate inflorescence ; the rosette pitchers are urceolate, the upper ones, when developed, are infundibuliform ; the rosette pitchers are always glandular upon the whole inner surface and the peristome is delicately ribbed. Paniculate inflorescences are, outside this group, only met with in the group of the Vulgatae.
.
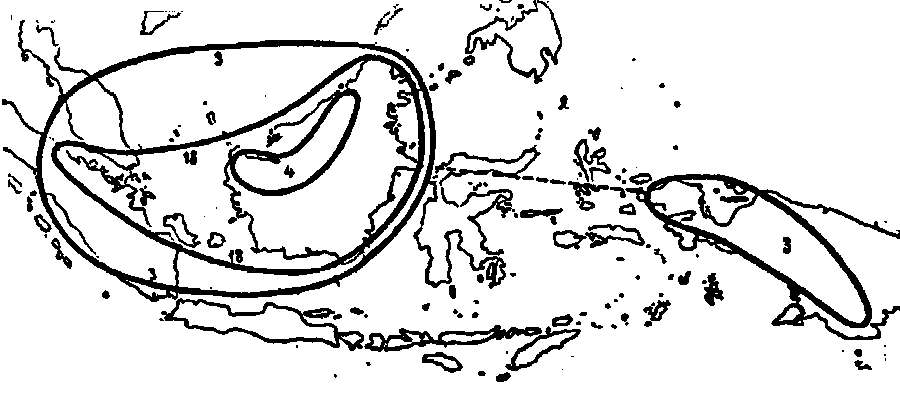 Fig. 34. The distribution of the Urceolatae ; 3. N. ampullaria ;
4. N. bicalcarata ; 18. N. Hookeriana.
Fig. 34. The distribution of the Urceolatae ; 3. N. ampullaria ;
4. N. bicalcarata ; 18. N. Hookeriana.
The above division is based by no means on phylogenetical considerations. To what degree it has a phylogenetical significance will be discussed further on.
As early as 1873, HOOKER F., in his monograph, gave a division into two subgenera as follows:
Sectio I. Aneurosperma. Semina ovoidea, testa inappendiculata. Fl. foem. Calycis tubus obconicus, limbo 3-4-fido. Ovarium obpyramidatum, vertice 3-4-lobo depresso, lobis apice intus stigmatosis.
Sectio II. Eunepenthes. Semina nucleo ovoideo, testa utrinque in caudam elongatam saepe longissimum capillarem producta. Fl. foem. Sepala 4, rarissime 3, patentia. Ovarium oblongum v. fusiforme, apice integro, stigmate discoideo latiore coronato.
To the first subgenus belongs only N. Pervillei, to the second all other species.
Not having seen N. Pervillei I have no critical opinion about this division, but I think it probable, that N. Pervillei has too much in common, with the other species I have inserted in the Vulgatae, that its subgeneric distinction from all other species of the genus not desirable. The character of the seed, which seems to be the principal argument for this separation, is not so important, since also in N. madagascariensis the appendages of the seed are very short. This seems to be also the opinion of MACFARLANE, who in his monograph has omitted the subgenera of HOOKER.
The further division of HOOKER in his monograph is undoubtedly meant as an artificial one and need not to be discussed here, the more so, as HOOKER has not named the divisions.
A second attempt to establish a natural subdivision is made by BECK in his "Monographische Skizze". Yet this division, indeed, is only seemingly a natural one. BECK follows HOOKER in distinguishing two subgenera and divides the subgenus Eunepenthes into three groups viz.: 1. Retiferae (l.c., p. 141), 2. Apruinosae (l.c., p. 141) and 3. Pruinosae (l.c., p. 183).
The first group, that of the Retiferae, comprises only one species, viz. N. Lowii, but as I have already pointed out, the aberrant characters of this species have not so much taxonomical value as they seem to have at first sight. The other two groups are distinguished by the presence or absence of a pruinose upper part in the inner side of the pitcher. Every botanist, however, who is acquainted with the polymorphy of Nepenthes, knows that this pruinose part is very differently developed in the different pitchers of most species. Though there are some species, which have no pruinose part in any of their pitchers, and some others which have it in all their pitchers, the greater part of them have it well developed in the lower ones and only little or not at all in the upper ones. Consequently it is impossible to base a classification of the species on this feature. For the distinction of these groups, BECK has principally considered the upper pitchers, in which the pruinose part is very differently developed in different species, and the consequence has been, that very closely related species have been separated, whereas very little related species have been put together. So among the Apruinosae we find a number of very nearly related species, e.g. N. Veitchii, N. maxima, N. celebica. some less related ones, such as N. Northiana and N. Rajah, but also N. trichocarpa, nearest related to N. gracilis, which justly has been inserted in the Pruinosae. Of N. ventricosa and N. Burkei, which are so nearly related that they must probably be considered as forms of a single species, we find the former among the Apruinosae, the latter among the Pruinosae. Among the Pruinosae were mark also N. stenophylla, closely related to N. maxima, N. spuria, synonymous with N. Northiana, and N. Boschiana, only little related to the rest of the Pruinosae. The groups of BECK evidently having no taxonomical value, it is needless to discuss the further subdivisions.
The division, given by MACFARLANE, though being a very good general synopsis, can not be considered as a natural division of the genus and is certainly not purposed to be so. Also this arrangement needs not to be discussed in detail, as MACFARLANE has not named the groups distinguished by him.
3. Phylogenetical.
As has already been pointed out at the beginning of the discussion on the systematic relationships, it is not possible to say, which family of plants may be considered as the nearest relation of the Nepenthaceae from a phylogenetical point of view. Accepting this, we may ask in the first place, where in the world Nepenthes may have had its point of origin. It is not at all necessary, that this has been in the actual area of distribution, and certainly not in one of the actual centers of evolution. It is permissible to seek the origin in regions that formerly had a favorable climate, but have it no longer.
Firstly we will define our position on the standpoint of the doctrine of the constancy of continents and oceans.
Concerning the total generic area, we have already observed that Nepenthes must be a relatively young genus, since it cannot be accepted that the outer boundaries of the range of the genus, especially those in the east, are impassable barriers to its spread. There the limit runs between New Guinea on one side and the Bismarck Archipelago and the Samoan Islands on the other. Since Nepenthes can live in New Caledonia, we may suppose, that it may still continue its dispersion over the islands north in which certainly have a more favorable climate.
The relative youth of Nepenthes appears also from the large number of species, which are found on one mountain only, viz. 25 out of 65. Of course, several of these species may, in the future, prove to be more widely spread or to be forms of other species, but it is, on the other hand, especially such local species, which have been discovered by the most recent expeditions to the interior of the greater islands of the Malay Archipelago, and therefore we may expect that their number will increase in the future. Such species cannot maintain their position during long periods ; they must spread or die out. If this line of thought be correct we must consider the regions with many local species as the centers of actual development, i.e. in the first place Borneo, in the second place Sumatra and New Guinea, in the third place Mindanao, the Malay Peninsula, Cochin China and Selébès. It is, however, not at all necessary to accept that it is in one of these regions that the genus has originated.
Asking, with what forms the genus has started its development, we are, of course inclined to consider the most simple forms as the most primitive, i.e. the Vulgatae. This agrees with the fact, that only the range of the Vulgatae is as large as that of the whole genus and that the most widely spread species, viz. N. mirabilis, belongs to this group. In this case it is permissible to apply WILLIS's principle of "Age and Area", as there is no reason to suppose, that the Vulgatae have in general much better means of dispersion than any of the other group.
Considering not from a phylogenetical standpoint the separate group distinguished by me, we may first ask, which forms of the Vulgatae must be considered as the most primitive. I have already observed, that we may distinguish between species with racemose and such with paniculate inflorescences. That in the synoptical list of the species on p. 405 I have placed those with panicles in the first column, was only due to the fact, that those with racemose inflorescences formed the connection with the following and most closely related groups. Now it is generally accepted, that the racemes of Nepenthes are only simplified panicles, and therefore we have good reason to suppose, that the species with paniculate inflorescences are in general the primitive ones. I am very well aware, that a reasoning like this is very dangerous and of little value, as nature, being able to form species with racemose inflorescences from such with paniculate ones, undoubtedly may be able to make also species with panicles from such with racemes. In general we have little idea, which metamorphoses occur more easily or with greater difficulty in nature, and this is the reason that phylogenetic considerations based on morphological differences are mostly of little value.
No more success have considerations, based on the Age and Area principle of WILLIS. I take the liberty to suppose, that this hypothesis and its theoretical consistencies are known to the reader and I must restrict myself to draw attention to the facts, that cannot be explained with the help of WILLIS's theories.
In the first place we can state the fact, that several species are isolated in the western extreme part of the total generic area. Looking first at N. madagascariensis we have to explain, where this species has originated, and how it has come to Madagascar. To give an answer to this question we must know in the first place, whether N. madagascariensis is a well defined species or not, i.e. whether N. madagascariensis, growing together with other species, remains separated from them, or intermixes with them, finally forming a mixed population, in which the original forms can no more be sharply distinguished. In the event, that it is only a form of another species, the origin is readily explicable, being caused by the climate of Madagascar, certainly differing from that of the country, where the other races of the same species occur. As I think it impossible, that such races gradually loose their possibility of intermixing with the other races of the same species (I am well aware that most botanists do not agree with me in this respect!), the origin of N. madagascariensis seems difficultly explicable, when we have to consider it as a separate species. And provisorily we are obliged to maintain it as such, since we do not know another species, of which it might be a form. Even the closely related N. Pervillei is undoubtedly a separate species, as it has sessile leaves and N. madagascariensis has petiolate ones, these two leaf forms never occurring together in any one of the other species, as far as I know. Another supposition is, that N. madagascariensis occurs somewhere in another country, where it grows together with other species. In the neighbourhood of Madagascar, however, there are no countries, In which this species may still be discovered. It is also improbable, that in Madagascar one of the other species will be discovered in future. So we have to suppose, that either N. madagascariensis has been derived in Madagascar from any other species now extinct, or that it has originated elsewhere, and that, after its migration, it has died out in its original birthplace.
This, however, leads to an unexpected consistency, since these considerations also apply to the other species of the western extreme part of the total generic area, viz. N. Pervillei, N. destillatoria and N. khasiana. For the explanation of the isolated situation of all these species dying out of several species should be supposed and this does not agree with the supposition, that Nepenthes is still spreading westwards and approaching Africa. On the contrary we have to accept, that Nepenthes has been more developed formerly in the western part of its present actual area.
Another case is extant in the eastern extreme part of the area of Nepenthes. Here N. Vieillardii is the only species that has reached New Caledonia and the Isle of Pines. The origin of this species, however, is easily explainable, since its occurrence in New Guinea, together with many other species, has been proved. Probably N. Vieillardii has originated in New Guinea, and by its greater hardiness it has succeeded in spreading farther eastwards than any other species.
If, bearing in mind WILLIS's theory of Age and Area, we seek the older species among those with the widest dispersion, we have to consider in the first place N. mirabilis, in the second place N. gracilis, N. Reinwardtiana, N. albo-marginata and N. tentaculata. Indeed all these species are very primitive ones. N. mirabilis seems not to be restrained by broad water such as the Straits of Malacca and the Straits of Makassar or the sea between the Moluccas. Certainly this is partly due to the fact, that N. mirabilis is rather indifferent regarding the elevation as well as regarding the temperature and the humidity of the habitat, partly also to its great age but I would not agree with WILLIS in his theory, in considering N. mirabilis as the ancestor of its genus.
Of the other Vulgatae already mentioned for their large area. N. gracilis, N. albo-marginata and N. Reinwardtiana are almost restricted to the Sunda-shelf, the first being found twice in Selébès, whereas N. tentaculata is common in all parts of Borneo and has been found once in Selébès. When discussing the distribution in general I have already supposed, that this has a geological cause, and I will try to explain this when discussing the phylogeny.
A very peculiar dispersion is that of N. alata, a species being found in Sumatra, the Malay Peninsula and the Philippines, but not in Borneo. If in future it is not discovered in the latter island, we will have to suppose either that it has originated polyphyletically or that it has died out in Borneo.
If they will later prove to be well defined species, N. paniculata, N. neoguineensis, N. tomoriana, N. papuana, N. trichocarpa, N. tobaica and N. Thorelli may be considered as young forms of an old group. None of these species gives reason to the supposition, that daintiness regarding the habitat is the cause of the limited dispersion, as is the case in so many species of the Regiae.
We now come to the Montanae, almost restricted to the mountains of the Malay Peninsula, Sumatra and Java. From these species, 3 occur in the Malay Peninsula, and 7 in Sumatra ; one of the latter has also been found in Java and perhaps in the extreme south of Borneo, this being also the species that is most similar to the Vulgatae. We may, therefore, suppose that the development of the Montanae has started in the mountains of Sumatra producing there 7 species, having spread on one side to the Malay Peninsula and having produced there 3 new species, whereas one of the species originated in Sumatra has succeeded in reaching Java, without producing new ones. This small phylogenetic success of this group in the southern part of its area may, at least partly, be ascribed to climatic conditions in Java, becoming less favourable towards the east. It is far from certain, whether N. gymnamphora, which is most similar to the Vulgatae, is the ancestor of the other species. Also it is questionable, whether the numerous intermediates between the species of this group are only rare forms, equivalent to the species from a phylogenetical point of view, or hybrids, originated secondarily from the species. Further investigations will probably throw light upon this question. As to the Geological period, in which the development of this group has started, we can say only, that this must have happened after the mountains of Sumatra and the Malay Peninsula had been formed, i.e. in the second half of the Tertiary period. The small area of distribution of the whole group and of most of its species, as well as the difference between the Sumatra and Malay Peninsula species is explained by the fact, that they are mountain plants, which never grow at low altitudes, and not disperse with help of the interjacent valleys nor of that of the periodically dry seas.
The group of the Nobiles is probably no phylogenetic unit, and therefore it has no value for phylogenetic discussions. The total area of the group is small and partly overlaps that of the Regiae, partly extends west of it.
I will not go so far as to assert, that the groups of the Regiae and of the Insignes have had an independent phylogenetic evolution, and I will, therefore, emphasize once more, that the distinction of these groups is not based upon phylogenetic argument. When we compare the areas of these two groups, we see, that the Insignes are a little more widely spread than the Regiae. The Regiae occur all over Borneo, whereas one species is spread all over the Moluccas and New Guinea, one species is only known from one locality in western New Guinea, and one species only from a single mountain in Mindanao. The Insignes, on the other hand, extend from Mindanao to western Sumatra on one side and to western New Guinea on the other, but they are not found in the Moluccas.
Paying attention firstly to the Regiae, it is obvious that for this group Borneo is the centre of development. Here not only the largest number of species occurs, but here we find also almost all local species. In the group of the Vulgatae only 3 out of 22 species had been found on one mountain, whereas of 15 Regiae the number of such species amounts to 11. Of these 11 there have been found 8 in Borneo, 1 in Mindanao and 2 in New Guinea. The remaining 4 species show a larger area of distribution, viz. N. stenophylla, N. Veitchii, N. Lowii and N. maxima, the latter having a very large area, viz. from western Borneo to eastern New Guinea. Now in this group it is undoubtedly wrong, to ascribe the limited distribution of most species to their relative youth only. We must also bear in mind, that most species are mountain plants, which migrate with great difficulty from one mountain summit to the other. I shall discuss the question of the means of distribution later. Yet it is remarkable that the widely spread N. maxima does not grow at low altitudes, the elevations recorded varying between 600 and 2100 m, whereas N. stenophylla, the second species as regards the area of its dispersion, seems to occur between 900 and 2667 m above the sea ; N. Veitchii on the contrary, though not a plant from the plain, seems to occur especially on lower mountains.
After all, this does not explain the wide area of N. maxima. Supposing N. maxima to have originated in Borneo, it has to be explained, why it has succeeded in migrating so far to the east and why it has not reached Sumatra, the former way being apparently much better explicable than the latter, which is only passable for very few species. lt is very improbable, that N. maxima might have originated in the Moluccas, where the conditions for Nepenthes seem to be generally unfavourable, or in New Guinea, as in this case all less wide spread allies should have originated from it after its arriving in Borneo. Also the supposition that the origin of N. maxima should have been quite independent from that of the other Regiae is unadmissable, several of the latter being so closely similar to N. maxima, that their distinction is often difficult.
Looking now at the species enumerated under the name of Insignes, we remark 10 species, 4 of which have been found on a single mountain only ; one, N. Merrilliana, on one or two localities ; one, N. insignis, on 2 localities ; whereas N. ventricosa and N. Burkei are spread over a great part of the Philippines and N. Rafflesiana all over the Sunda-shelf. This is almost the same as we have seen for the Regiae.
If considering together the Regiae and the ns the same, we then have 25 species, 15 of which have been found on a single mountain only, 3 have been found in 2 localities, 5 have a larger area of distribution and 2 are rather widely spread. If one would conclude from this, that N. maxima and N. Rafflesiana have to been reckoned among the older species of these groups, I could concur in his opinion. When, however, one would conclude, that they were the species, from which all others have taken their origin, I should remark, that this does not agree with the morphological characters. So we see that in both groups the manner of development of the species is unknown.
From the groups distinguished by me, there still remains that of the Urceolatae, rather uninterest from a phylogenetical point of view, as it certainly is no phylogenetical unit. N. ampullaria has a remarkable disjunct area of dispersion, consisting of two parts: the Sunda-shelf and New Guinea. The morphology of this species gives no indication for an explanation of its genesis. Perhaps it is a very old species having originated from the Vulgatae quite separately from others of N. bicalcarata the same maybe said, though its area is much more common, being limited to the mountains of northwestern Borneo. N. Hookeriana may remain out of consideration, as it is probably a hybrid of N. ampullaria and N. Rafflesiana.
In the above, certain regularities in the areas of dispersion have been stated, that reappear in different groups, and which require an explanation.
In the first place we have seen, that the species are not regularly spread over the total area of the genus, nor concentrated round one centre. We observe several centres of development, especially the mountains of Borneo, Sumatra, the Malay Peninsula, Farther India, Mindanao and New Guinea. This has to be ascribed to the fact, that mountainous regions are especially favourable for the development of this genus. Only few species are typical lowland plants and even these can grow in the mountains. Most of them, however, are plants of the mountains, partly occurring also in hilly land, partly confined to high summits.
Two other regularities concerning the areas can be explained. In the first place the fact, that in 3 of the 6 groups distinguished. there are species, whose area coincides fairly well with the Sunda-shelf. Undoubtedly this has in first line a geological cause. The Sunda-shelf is actually a group of islands, separated by shallow seas, but these seas, have alternatively been dry and flooded in the last geological periods. For species, which can grow in the lower hilly land or even in the plains, this must have provided a fair opportunity of migrating from one island to the other. For mountain species, however, it can not have had any significance. This agrees with the facts observed: N. gracilis, N. Reinwardtiana and N. albo-marginata in the group of the Vulgatae, N. Rafflesiana among the Insignes, N. ampullaria among the Urceolatae, show a distribution that seems to be influenced by the geological processes mentioned above, and all these are species, which can grow in the plains.
In the second place we have stated the fact, that in all of the 6 groups there are species, the dispersion of which is restricted to the mountains of central and northwestern Borneo. We may readily suppose, that they have originated in these regions, but this does not explain, why they have not succeeded in reaching the mountains of other islands. The cause is probably analogous to that of the restriction to the Sunda-shelf. The mountains of central and north-western Borneo have never been flooded in the last geological periods, probably never after their origin, and they are, therefore, favourable centre for the development of the more sensitive species. In the times that the seas of the Sunda-shelf were dry, these species, though becoming perhaps more widely spread, had no opportunity of using the lowland connections for their migrating to other islands and when the sea level rose again, they were driven back to their country of origin. Among the Nobiles it is N. hirsuta, among the Regiae it is N. Veitchii, N. stenophylla, N. Lowii, among the Urceolatae it is N. bicalcarata, which behave in this manner.
We generally accept, that the seeds like those of Nepenthes are dispersed by the wind and that, by their extreme lightness, relatively little time should suffice for a transport even over great distances. From the above, however, we may conclude, that the transport of the seeds over greater distances is not so easy, as most species are hindered in their dispersion by relatively small seas and valleys between mountains. In general this may be explained by the fact, that most Nepenthes are forest plants, and that in the forest the wind certainly will transport the seeds of Nepenthes over small distances, but undoubtedly not from one mountain to the other. The seeds of the species, that grow on barren mountain tops, will be transported over greater distances, but in general they will arrive on the other mountain at much lower elevation where the species cannot grow. Only those species which grow as well on open mountain tops as in the forest of lower regions may have more use of the wind. Such species are N. gymnamphora, N. maxima, N. mirabilis, N. Vieillardii and really these species have spread over greater distances or have crossed valleys and broader seas.
So we see, that much of the regularities in the distribution of Nepenthes can be explained, but that much more remains incomprehensible.
Before passing to the summary of the results, I will consider the previously discussed facts once more in the light of WEGENER's theory on the origin of the continents and oceans, as it seems to me that this theory throws more light on the phylogeny of this genus.
I will begin with drawing attention to the fact, that Nepenthes occurs in Madagascar, but not on the mainland of Africa. This fact is stated for many plants and animals and is generally explained by supposing, that formerly there was a land connection between India and Madagascar over the Maldive Islands, the Seychelles and the Amirante Islands. WEGENER explains this by the hypothesis, that formerly Africa, Madagascar and India have been parts of one continent, broken into parts, that floated away in different directions. In the third edition of his paper we read on p. 2:
"Ebenso wird angenommen, dass Antarktika, Australien und Vorderindien bis zum Beginn der Jurazeit unmittelbar nebea Südafrika gelegen und mit diesem und Südamerika ein einziges grosses-wenn auch teilweise von Flachsee überschwemmtes-Kontinentalgebiet gebildet haben, das im Laufe von Jura, Kreide und Tertiär durch Zerspaitung in Einzelschollen zerfiel, die nach allen Seiten auseinandertrifteten. Unsere in Fig. 1 und 2 wiedergegebenen drei Erdkarten für das Jung-Karbon, Eozän und Alt-Quartär zeigen diesen Entwicklungsang. Bei Vorderindien händelt es sich dabei um einen etwas abweichenden Vorgang: Es war ursprünglich durch ein langes, freilich meist von Flachsee bedecktes Schollenstück mit dem asiatischen Kontinent verbunden. Nach der Abtrennung Vorderindiens einerseits von Australien (in der älteren Jurazeit), andererseits von Madagaskar (an der Wende von Kreide und Tertiär) wurde dies lange Verbindungsstück durch fortschreitende Annäherung Vorderindiens an Asien immer mehr zusammengefalten, und ruht heute in den gewaltigsten Gebirgsfaltet welche unsere Erde trägt, dem Himalaja und den zahlreichen weiteren Faitenzügen Hochasiens."
And 1. c. p. 59:
"Die biologischen Beziehungen zwischen Dekan und Madagaskar, angeblich über ein versunkenes "Lemurien" hinweg, sind so allbekannt, dass wit uns hier mit einem Hinweis auf unsere Fig. 15 und auf die Zusammenstellung bei Arldt begnügen wollen. Gerade hier zeigt sich wieder der Vorzug der Verschiebungstheorie, da die beiden Teile in ihrer heutigen Lage einen bedeutenden Breitenunterschied besitzen und nur deshalb ähnliches Klima haben und ähnliche Formen beherbergen können, weil der Aequator zwischen ihnen liegt."
.
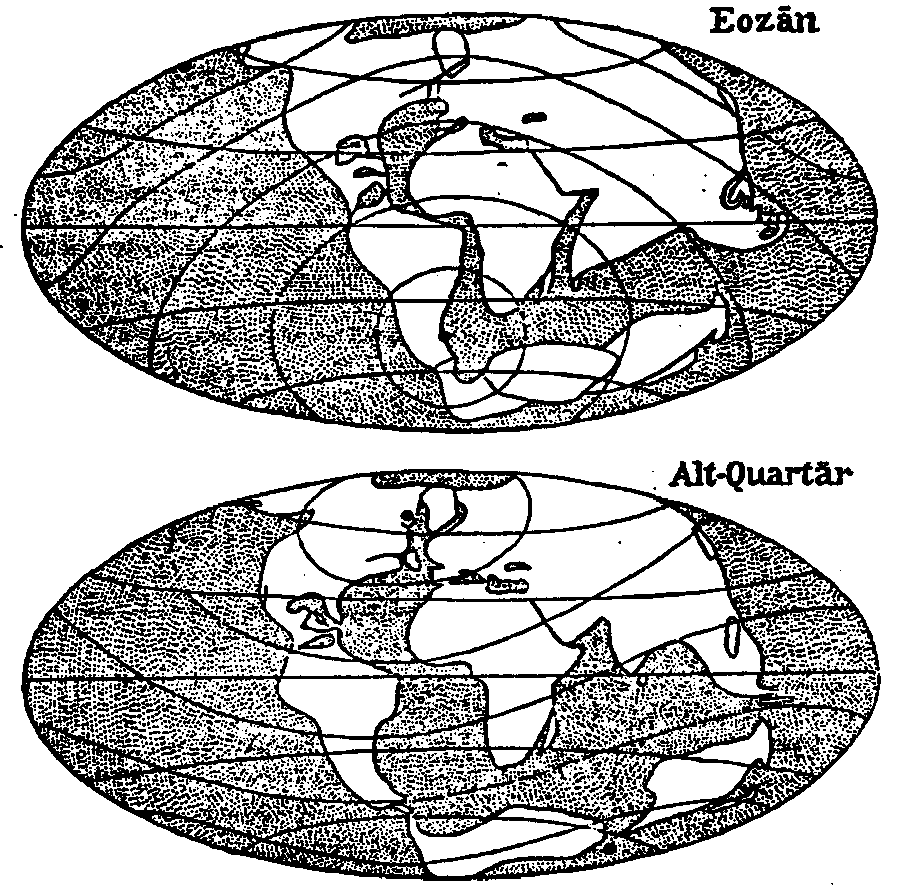 Fig. 35. The situation of the continents in the Eocene and in the beginning of
the Quartary period, according to WEGENER (after WEGENER and IRMSCHER).
Fig. 35. The situation of the continents in the Eocene and in the beginning of
the Quartary period, according to WEGENER (after WEGENER and IRMSCHER).
Looking at some of the maps WEGENER gives (fig. 35), we see that Madagascar, according to his theory, originally was situated between Africa and the southern part of India, near Ceylon. The Seychelles have to be considered as small fragments of land, remained behind from Africa and Madagascar. WEGENER supposes (i.c. p. 40) that they have emerged from the ocean as parts of the African continent, left behind on the bottom of the sea, but this would not explain the occurrence of Nepenthes in the Seychelles.
From this we may conclude, that in the beginning of the Tertiary era Nepenthes already existed, and provisionally we can think the genus to have originated during the last part of the Cretaceous. When the mainland of Africa was separated from Asia, Nepenthes had not yet reached the former. When Madagascar and the Seychelles floated away from India, they took with them only the primitive forms existing at that time. This also elucidates, how it is possible, that some species might have crossed the large seas between India and the Seychelles and between the Seychelles and Madagascar, while they have not crossed the straits between Madagascar and the mainland of Africa. About the latter we read l.c. p. 42:
"Madagaskar besteht wie das benachbarte Afrika aus einer Tafel gefalteten Gneises mit nordöstiicher Streichrichtung. An der Abrisslinie sind beiderseits identische marine Sedimente abgelagert, welche andeuten, dass seit der Trias beide Länder durch einen überschwemmten Grabenbruch getrennt waren, was auch die madagassische Landfauna verlangt. Aber noch in der Mitte der Tertiärzeit, als Indien bereits abgerückt war, sind nach Lemoine zwei Tiere, der Potamochoerus und der Hippopotamus, von Afrika eingewandert, die, wie Lemoine meint höchstens einen Meeresarm von 3O km Breite dürchschwimmen konnten, während jetzt der Kanal von Mozambique gut 400 km breit ist. Erst nach dieser Zeit kann sich also die madagassische Scholle auch untermeerisch von Afrika losgerissen haben, wodurch sich der weite Vorsprung erklärt, den Vorder-Indien in der Verschiebung nach Nordosten gegenüber Madagaskar bekommen hat."
From this it is obvious, that the species of Nepenthes, that inhabitated Madagascar at the time of its separation from the Asiatic continent, have not been able to cross a strait, that in the beginning was at most 30 km broad ; neither may we expect it crossing a much broader strait in future.
These considerations do not tell us anything about the occurrence of Nepenthes in the eastern part of its present area of dispersion. According to WEGENER New Guinea has always been a part of the Australian shelf and together with this it has been separated from Asia and it has first floated southwards, than turned with its eastern part to the north, till New Guinea reached and penetrated the Malay Archipelago, which always had been a part of the Asiatic land-mass. If, during its separation from Asia, Australia has possessed any Nepenthes, these species must have died out during the time that Australia floated to the south, the more so, since about that time the South Pole was situated much more to the North (see fig. 35). On this point, WEGENER says (l.c., p. 46-47):
"Von diesen letzten Bewegungen Australiens erzählt uns namentlich die Tiefenkarte der Umgebung von Neuguinea mancherlei Einzelheiten. Die grosse australische Scholle drängt sich, wie Fig. 12 schematisch erläutert, mit ihrem ambossartig verdickten votderen Ende, dem zu einem hohen jugendlichen Gebirge aufgefalteten Neuguinea, von Südosten kommend, zwischen die früher vermutlich geschlossenen Ketten der südlichsten Sunda-Inseln und des Bismarck-Archipels ................ Diese Kette wird weiterhin in einer ähnlichen energischen Spirale bis Buru zurückgebogen. Und eine sehr interessante Ergänzung zu diesem Vorgang sieht man auf der Ostseite Neuguineas ; Von Südosten kommend, hat dies die Inseln des Bismarck-Archipels gestreift, dabei die Insel Neupommern an ihrem früheren Südostende erfasst and mit sich geschleppt, die lange Insel um mehr als 90o herumdrehend und halbkreistörmig biegend".
According to the maps given by WEGENER, the penetration of New Guinea into the Malay Archipelago has taken place in the Diluvial period. All the forms of Nepenthes, occurring now in this island, must consequently have penetrated from the West, or have originated there. This seems improbable as, according to WEGENER, the Louisiade Archipelago with N. mirabilis, and New Caledonia with N. Vieillardii, have remained behind from New Guinea during its voyage to the Malay Archipelago, from which we might conclude, that the 2 species mentioned should originate from the Australian land mass. As we have already seen, this is impossible. Consequently, we are obliged to accept, that the islands of the Louisiade Archipelago and New Caledonia with the neighbouring islands, have first been connected with New Guinea and have been separated and floated back afterwards. In this respect a study of the New Caledonia plants, which only can have migrated from the Malay Archipelago (such as Nepenthes) should be of great interest.
Summarizing the results of the foregoing discussions, we come to the following short phylogenetic history of the genus.
Nepenthes has originated in the Asiatic continent during the Cretaceous period, in the regions, that now form the Himalaya and India, but about that time were situated more southwards, and partly south, partly north of the then equator (cf. fig. 35). As in the beginning of the Tertiary period, Africa became separated from India, the genus had not yet reached this part of the world. When later Madagascar was separated from India, that island possessed at least one species of the paniculate Vulgatae, of which N. madagascariensis is the descendant. The same was the case in he Seychelles and Ceylon. The racemose Vulgatae perhaps existed already or were in statu nascendi during the travelling of the genus to the east. As the landmasses, which were then the centre of evolution, were transformed into India as it is at present, the conditions for Nepenthes grew gradually less favourable, whereas the landmasses, which now form the Malay Archipelago in the same time approached the equator and became more favorable regarding to the climate. In this way species died out in the western part of the total generic area and the center of evolution moved gradually to the east. The first forms, that penetrated southern Asia were paniculate as well as racemose Vulgatae, but it can not be stated, whether the racemose or the paniculate were the pioneers. In the Archipelago, parts of the genus became isolated, and this furthered the origin of separate groups. Thus the Montanae took their origin in the mountains of Sumatra and the Malay Peninsula, one species of the group, N. gymnamphora, migrating afterwards to Java and to southern Borneo. As this group contains mountain forms only, it could not reach the centre of Borneo from Sumatra directly. With, or partly without, help of temporarily dry seas, the Vulgatae reached not only all the islands of the Sunda-shelf, but also the Moluccas and the Philippines. In Borneo the conditions were extremely favorable for the development of new forms, which now are called the Regiae and the Insignes, strange and beautiful, but very sensitive regarding to climatic alterations. As a result of this, these groups are remarkable for the many species that have not spread farther than one mountain summit, where it probably originated. Both groups reached the Philippines and New Guinea in few species. When New Guinea approached the Molluccas, only the Vulgatae and N. maxima had hardly reached these islands, which is the cause, that still now no Nepenthes occur east of New Guinea. Rapidly New Guinea was occupied by Vulgatae, both paniculate and racemose, and by N. maxima. Though this must have happened not earlier than in the Diluvial period, new forms have already originated in the western part of these islands, there the conditions for the growth of Nepenthes being most favourable. One of the species, more hardy than other ones, has reached the Louisiade Archipelago, another, likewise very hardy, has reached New Caledonia and the isle of Pines, probably at a time when these islands were connected with, or situated much nearer to, New Guinea. About the origin of the Nobiles and the Urceolatae nothing is known. Perhaps the Urceolatae originated separately from the paniculate Vulgatae.
________
ADDITION
The receipt of the Nepenthes collected by Prof. HANS WINKLER in Western Borneo when this revision had already been printed for the greater part, makes desirable the following addition.
Ad 2. Nepenthes albo-marginata LOBB (p. 262).
BORNEO. Res. Western Division: on the Sg. Bika, in marshy forest, 4 II 1925, HANS WINKLER 1590 (0) ; on the Bt. Tiloeng, 800 m, 8 II 1925, HANS WINKLER 1491 (0).
N. albo-marginata was already known from the upper Kapoeas valley.
Ad 12. Nepenthes ephippiata DANS. (p. 286).
Stems climbing often up to the tops of the trees. Lower pitchers of the climbing stems rather abruptly originating and shortly incurved from the hanging tendril, sharply triangular in the curved part, broadly rounded at the base, about 15 cm high, 8 cm wide in the lower part, slightly but distinctly narrowed at half the height (reckoned from the back side), slightly but distinctly infundibuliform towards the mouth, with 2 prominent ribs over the whole length bearing a fringed wing rudiment at some distance from the peristome ; mouth oblique, acute towards the lid ; peristome in front cylindrical, about 3 mm broad, with ribs l to 1 1/2 mm apart, towards the lid flattened and broader, with ribs gradually coarser, about 7 mm broad and 3 to 4 mm apart near the lid, the inner part slightly developed in the front part but not toothed, wholly failing towards the lid ; spur none ; lid ovate, rounded at the apex deeply cordate at the base, about 15 cm long, 11 cm broad, wholly vaulted, the inner surface with many hundreds of appendages in the basal half with exception of the marginal part about 2 cm broad, the appendages densely set and laterally flattened towards the thickened basal part of the midrib, less densely set and more nipple-shaped towards the margin, the whole inner surface, with exception of a marginal part about l l/2 cm broad, rather densely glandular with thickened and deepened
.
(HANS WINKLER 1023)
 Fig. 36. Nepenthes ephippiata, 1/2 x.
Fig. 36. Nepenthes ephippiata, 1/2 x.
glands covered by the thickened rim and having a very narrow mouth ; inner surface of the pitcher wholly glandular, the glands in the lower part large and flat, not overarched and only separated by narrow ribs forming irregular polygons, these glands growing smaller towards the narrowed part of the pitcher, rather abruptly replaced by smaller normally overarched glands above the narrowed part, growing very small round and deepened towards the peristome, the uppermost ones again rounder and larger and resembling the very distinct marginal glands of the peristome ; about 200 to 250 glands on 1 cm2 at the bottom of the pitchers, about 400 to 600 above the narrowed part, about 400 to 500 towards the peristome. Male inflorescence a robust raceme, the peduncle about 17 cm long, flattened, about 8 mm broad in the basal part, 6 to 7 mm towards the raceme, the axis about 30 cm tong, gradually attenuate towards the tip, grooved, the pedicels densely set, the lowermost ones about 22 mm long, inserted about 5 mm above the base, the upper ones gradually a little shorter, the outermost ones about 10 mm long, all of them 2-flowered, without bract. Tepals of the male flowers elliptical, about 3 1/2 to 4 1/2 mm long, rounded at the apex. Staminal column about 5 mm long, the anthers included, which are situated in 1 whorl and an apical group. Indumentum of the male inflorescence like that of the female one, the staminal column shortly densely stellate-tomentose at the base, glabrous towards the anthers. Colour of the living plant: leaf blade shining-green, midrib yellow above, light-green below ; tendril light-green, pitcher light-green, somewhat reddish near the peristome, the 2 longitudinal ribs and peristome light-red ; inner surface dark-red andshining above the narrowed part, green with red points below the narrowed part ; lid green above, the nerves red towards the margin, in older pitchers the whole upper surface red ; lower surface shining yellowish-green near the insertion, the rest green with a red margin about 5 mm broad and with red appendages and translucent-white glands with red rims ; hairs of the leaf and the tendril brown.
BORNEO. Res. Western Division: on the Bt. Raja. 1900 m, 22 XII 1924, HANS WINKLER 1023 (m).
Though the type specimen, described on p. 286-288, bears no pitchers and of the generative organs only female flowers and fruits, whereas the plant collected by WINKLER bears pitchers and male flowers, I do not doubt, whether both belong to the same species. The stem fragment of WINKLER's plant shows the same peculiar structure as that of the type specimen, the indumentum is the same, the leaves show only slight differences, and also the nervation of the leaves of WINKLER's plant is as described on p. 286 and figured in fig. 5. The only pitcher rudiment of the type specimen shows already the peculiar base and the relatively large lid, so typical for WINKLER's plant. The structure of the female and the male inflorescence is the same, the colour of the herbarium specimens shows hardly any difference. Slight differences however are to be stated: WINKLER's plant has leaves somewhat more acute at the apex and at the base, the pennate nerves a little more distinctly forming partial longitudinal nerves.
The pitcher shows, that I have justly placed N. ephippiata into the section of the Regiae: it even belongs among the strangest forms of the genus. In several respects the pitchers are intermediate between those of N. Lowii and N. Rajah. Confer e.g. the characters of the lid and the glands of the inner surface of the pitchers. The appendages agree with those of N. Lowii as to the number and the place, but those placed towards the basal part of the midrib show the same shape as the single appendage on the lid of several other Regiae.
The description has been made after the only pitcher received at Buitenzorg. The label bears, besides on ample description of the colours, e.g. the following remark: "Sehr feste lederartige Beschaffenheit. Wenn man mit Holz gegen den Becher schlägt, klingt es wie wenn man einen Topf schlägt. Bei vielen Kannen ist der Deckel so stell aufgerichtet, dass er nicht als Regenschutz dienen kann". Yet the pitcher is by far not so hard as that of N. Lowii, which may be called really woody.
The second habitat is situated not far from the first ; yet the distance is so great, that a larger distribution in the mountains of Central Borneo is probable.
Ad 22. Nepenthes leptochila DANS. (p. 319).
BORNEO. Res. Western Division: on the Bt. Mehipit, 500 m, 27 XII 1924, HANS WINKLER 1100 (0) ; between Bidang Menabai & Bt. Mehipit, 600 m, 27 XII 1924, HANS WINKLER 1101 (0).
The specimens, collected by WINKLER, bear, like the type specimen, no flowers, and give no occasion for a completion of the description. The leaves of one of the stems are somewhat longer and more rounded at the apex and bear 4 or 5 longitudinal nerves on each side. The peristome of some pitchers is as narrow as is described an p. 321 ; of other pitchers, however, it is up to 3 mm broad. It is peculiar, that the new habitat is so far remote from the first and this makes probable a larger distribution of the species.
Ad 44. Nepenthes tentaculata HOOK. F. (p. 379).
BORNEO. Res. Western Division: Bt. Raja, 1400 m, 21 XII 1924, HANS WINKLER 1004 (m) ; ibidem 24 XII 1924, idem 1053 (0).
N. tentaculata was the only species known from the Bt. Raja up to the present.
___________